- Ramp protocol изменения мембранного потенциала
- Методика фиксации мембранного потенциала (МП) и регистрации трансмембранных токов (Iм)
- ОБЩИЙ МЕМБРАННЫЙ ТОК I m
- Ионные токи, зарегистрированные методом фиксации потенциала
- Фиксация потенциала на гигантском аксоне кальмара
- Избирательное блокирование натриевых
- МЕТОД patch- clamp и его
- Регистрация мембранного потенциала
- Мембранный потенциал
- Содержание
- Физический состав
- Измерение
- Физиологические ценности
- Изменения потенциала как сигналы
- Основы создания
- распространение
- Равновесный потенциал
- Общий мембранный потенциал
- Поддержание постоянной концентрации
- Получение через свободную энтальпию
- Химический потенциал — без учета разности потенциалов
- Электрохимический потенциал
- Уравнение Нернста
- Уравнение Гольдмана-Ходжкина-Каца
Ramp protocol изменения мембранного потенциала
До сих пор мы рассматривали токи и сдвиги проводимости всей мембраны при ее деполяризации. Несколько лет назад был разработан метод регистрации токов в микроучастках мембраны диаметром примерно 1 мкм, который позволяет идентифицировать молекулярные реакции одиночных каналов на основе зависимостей ионных токов от потенциала и времени. Рис. 2.11 иллюстрирует принцип локальной фиксации потенциала («patch clamp«) [12, 24]. Стеклянная микропипетка, диаметр кончика которой меньше 1 мкм, подводится к клетке вплоть до контакта с мембраной, и когда через пипетку подается отрицательное давление, пипетка обычно закупоривается участком мембраны; электрическое сопротивление между пипеткой и внеклеточным раствором возрастает скачком более чем до 1 ГОм (109 Ом). В результате микроучасток мембраны электрически изолируется от остальной мембраны. Канал пипетки соединен с усилителем обратной связи, который обеспечивает регулирующую цепь для поддержания потенциала пипетки на заданном уровне. Ток, необходимый для стабилизации потенциала-«ток фиксации»-точно соответствует току, протекающему в каждый момент через микроучасток мембраны. Командный потенциал усилителя можно устанавливать произвольно, так что регистрация токов через микроучасток мембраны может осуществляться при различных мембранных потенциалах или после ступенчатых сдвигов потенциала.
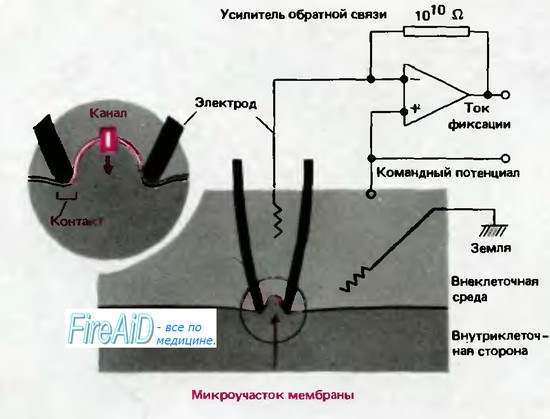 Рис. 2.11. Схема локальной фиксации мембранного потенциала («пэтч-кламп»). Изображен продольный срез через регистрирующую микропипетку (обозначена черным цветом) с диаметром контактирующего с мембраной кончика
Рис. 2.11. Схема локальной фиксации мембранного потенциала («пэтч-кламп»). Изображен продольный срез через регистрирующую микропипетку (обозначена черным цветом) с диаметром контактирующего с мембраной кончика
1 мкм. Если кончик электрода абсолютно чист и поверхность клетки освобождена от волокон соединительной ткани, то при подаче через пипетку отрицательного давления образуется тесный контакт, который создает электрическую изоляцию каналов находящегося в кончике пипетки микроучастка мембраны от остальной мембраны клетки (вставка). Таким способом можно регистрировать токи каналов с помощью усилителя обратной связи, соединенного с раствором электролита в пипетке (по [12, 24] с изменениями)
Гигаомный контакт между пипеткой и мембраной настолько прочен, что после отведения пипетки микроучасток мембраны часто отрывается от клетки, оставаясь прикрепленным к кончику пипетки. В этом случае регистрацию можно производить в микроучастке мембраны, отделенном от клетки, причем цитоплазматическая поверхность этого участка может омываться любым нужным раствором. Путем искусных манипуляций микроучасток мембраны можно даже перевернуть на пипетке наружной стороной мембраны наружу. Тогда цитоплазматическую поверхность можно орошать раствором в пипетке, который должен примерно соответствовать внутриклеточной среде, а на наружную поверхность могут воздействовать растворы различного состава; такая конфигурация «наружной стороной наружу» („outside-out») очень полезна для тестирования реакций каналов мембраны на изменения состава внеклеточной среды, на медиаторы или на фармакологические средства внеклеточного действия. Достаточно прочный контакт между участком мембраны и кончиком пипетки может быть достигнут только при абсолютной чистоте стекла пипетки и мембраны. Образованию контакта могут мешать волокна соединительной ткани, которые обычно приходится удалять путем обработки мембраны такими ферментами, как коллагеназа [12].
Источник
Методика фиксации мембранного потенциала (МП) и регистрации трансмембранных токов (Iм)
Ус — усилитель, реагирующий выходным током на разность между задаваемым «извне» потенциалом Е и МП . В силу конструкции системы ток I этого усилителя, проходя через сопротивление мембраны ( Rм ) изменяет МП так, что достигается равенство между МП и Е. При достаточном коэффициенте усиления усилителя и быстродействии системы МП практически фиксируется на уровне Е. При снижении Е и вслед за ним МП до КУД или более в мембране нервного волокна (кальмара) открываются потенциалозависимые натриевые и калиевые каналы, что порождает трансмембранные токи, которые и регистрируются на фоне поддерживаемого сниженного МП.


ОБЩИЙ МЕМБРАННЫЙ ТОК I m
I m C dV dt I i
Емкостной Общий ионный ток ток
I i I Na I K I L ,

Ионные токи, зарегистрированные методом фиксации потенциала
А — ток, протекающий через мембрану (синяя кривая) при смещении потенциала до 0 мВ относительно поддерживаемого потенциала, равного -60 мВ (поддерживаемый и стимулирующий ток выделен красным цветом). Б — разделение мембранного тока (I m ) на калиевую и натриевую
компоненты : 1 — аксон находится в физиологическом растворе, I = I Na + I K ;
2 — натрий заменен на холин, I = I K ; 3 — разность между 1 и 2, I = I Na .
Отклонение кривой вниз соответствует входящему току, а вверх соответствует выходящему току. Поддерживаемый потенциал мембраны клетки и его смещение обозначены красной кривой.

Фиксация потенциала на гигантском аксоне кальмара
А — смещения мембранного потенциала во времени
Б — ток через мембрану , регистрируемый одновременно со смещением потенциала. Показаны только смещения потенциала в положительную область от уровня поддерживаемого потенциала, равного -60 мВ (например,
В — вольтамперные характеристики , полученные в
результате экспериментов с фиксацией потенциала. По оси абсцисс — смещения мембранного потенциала относительно поддерживаемого потенциала (в данном случае потенциала покоя); по оси ординат — изменения входящего Na + -тока (фиолетовая кривая) и выходящего К + -тока (коричневая кривая)

Избирательное блокирование натриевых
и калиевых каналов с помощью тетродотоксина

МЕТОД patch- clamp и его
конфигурации для измерения токов через одиночные каналы

Na + -ток через одиночный Na + — канал в мышечной клетке мыши. А — Регистрация методом patch- clamp в конфигурации cell-attached
одиночных ионных каналов при смещениях мембранного потенциала от -80 до -40 мВ. Открытое состояние Na + -каналов представлено в виде смещения нулевой линии вниз, т.е. через канал течет входящий Na + -ток. Б — Регистрация методом patch- clamp в конфигурации outside-out
одиночных ионных каналов при смещениях мембранного потенциала от -100 до -40 мВ

Na + -токи, зарегистрированные в конфигурации whole-cell у электровозбудимых клеток при различных величинах смещения мембранного потенциала относительно поддерживаемого потенциала. K + -каналы были ингибированы Cs, тетраэтиламмонием или 4-аминопиридином.
А — Na + -токи, зарегистрированные у разных электровозбудимых клеток
Б — Na + -токи, зарегистрированные при разных ступеньках относительно поддерживаемого потенциала. Величина поддерживаемого потенциала E h равна -90 мВ.
Величины ступенек тестирующих потенциалов указаны на рисунке. В — вольтамперные характеристики,
построенные по пиковым значениям
(о) и по стационарным значениям (∆)Na + -токов.
Источник
Регистрация мембранного потенциала




![]()
![]()
В настоящее время наиболее часто для регистрации мембранного потенциала покоя используют стеклянные микроэлектроды. Первое их применение Лингом и Джерардом в 1949 году было не менее важным событием, чем введение пэтч-кламп метода три десятилетия спустя. Этот метод, кроме точного измерения мембранного потенциала покоя клетки, обеспечил регистрацию потенциалов действия, а также синаптических сигналов в мышечных волокнах и нейронах.
Метод внутриклеточной микроэлектродной регистрации проиллюстрирован на рис.14 Б. Острый стеклянный микроэлектрод (микропипетка), диаметр кончика которого не превышает 0.5 мкм, заполненный концентрированным солевым раствором (например, KCI), подводится к клетке, второй электрод помещается во внеклеточную жидкость (рис.14 Б1). Оба электрода присоединяются к усилителю и осциллографу для регистрации потенциала. В этом случае осциллограф регистрирует отсутствие какого-либо потенциала, поскольку оба электрода находятся снаружи клетки. В момент прокалывания микроэлектродом клеточной мембраны, приводящего к проникновению его в клеточную цитоплазму, осциллограф регистрирует появление отрицательного потенциала, соответствующего мембранному потенциалу покоя (рис. 14 Б 2). При удачном проникновении в клетку мембрана обхватывает внешнюю поверхность пипетки, благодаря чему цитоплазма остается изолированной от внеклеточной жидкости. Мембранный потенциал покоя широко колеблется в различных клетках (от -5 до -100 мВ). Наибольшие значения мембранного потенциала покоя зарегистрированы в возбудимых клетках: нервных, мышечных и секреторных, в которых его величина составляет от -60 до -90 мВ.
Мембранный потенциал может быть искусственно изменен при пропускании электрического тока через мембрану. Для иллюстрации возьмем вторую пару аналогичных электродов, подсоединенных к источнику тока, один из которых введем в клетку (рис. 15). При включении источника тока на короткое время через мембрану клетки будет течь электрический ток, несущий определенные заряды. Если ток имеет выходящее направление (внутриклеточный токовый электрод положителен по отношению к внеклеточному), то внутренняя поверхность мембраны становится более позитивной, а наружная — более негативной (рис. 15А). Это приведет к уменьшению зарядов на мембране и снижению мембранного потенциала. Уменьшение мембранного потенциала носит название деполяризации. Кратковременная деполяризация, возникающая при слабых толчках тока, называется электротоническим потенциалом, величина которого зависит от силы тока. В отличие от толчков тока, которые его вызывают, электротонический потенциал имеет медленное нарастание и медленный спад, что связано наличием в мембране сопротивления и емкости (RC цепочки). Изменение направления пропускаемого тока (внутриклеточный электрод отрицателен по сравнению с внеклеточным) ведет к увеличению отрицательного заряда на внутренней поверхности мембраны, и мембранный потенциал увеличивается (рис. 15 Б). Это увеличение абсолютного значения мембранного потенциала обозначается как гиперполяризация. Величина гиперполяризации также зависит от силы пропускаемого тока.
В вышеприведенных экспериментах сдвиг мембранного потенциала происходит из-за перераспределения зарядов на мембране при пропускании электрического тока. В естественных условиях очень часто деполяризация или гиперполяризация мембраны возбудимых клеток возникает при открытии потенциал-активируемых или лиганд-активируемых ионных каналов. Так, открытие натриевых или кальциевых каналов и поступление положительно
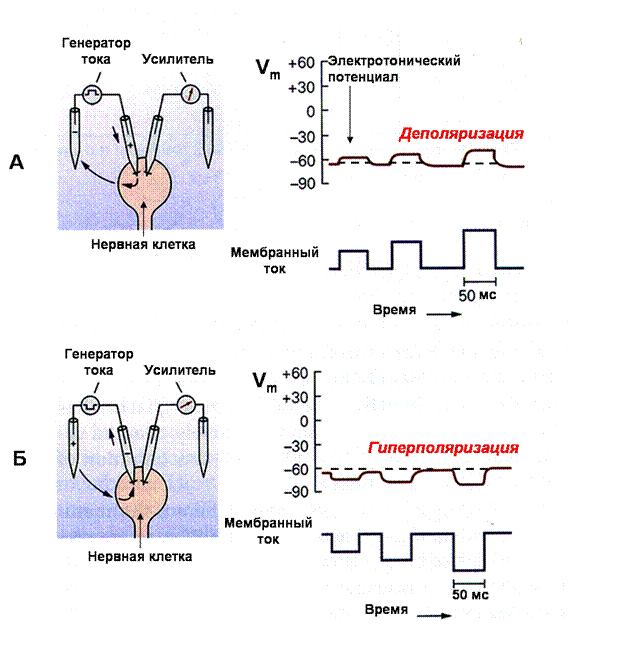
Рис 15. Изменения мембранного потенциала при пропускании электрического тока
(А) Один внутриклеточный микроэлектрод введен в клетку и регистрирует мембранный потенциал покоя, как показано на рис.14Б. Другой внутриклеточный микроэлектрод также введен в клетку и служит для пропускания слабых кратковременных толчков тока от генератора. Если пропускаемый ток имеет выходящее направление, то за счет перераспределения зарядов на мембране мембранный потенциал клетки уменьшается (деполяризация). Величина деполяризации зависит от силы тока.

(Б) Аналогичная схема, но пропускаемый ток имеет входящее направление. В этом случае мембранный потенциал увеличивается (гиперполяризация).
заряженных ионов в клетку будет приводить к деполяризации. Открытие калиевых каналов и выход положительно заряженных ионов калия из клетки вызовет гиперполяризацию. Открытие хлорных каналов нервных клеток обычно сопровождается поступлением отрицательно заряженных ионов хлора в цитоплазму и вызывает гиперполяризацию мембраны.
Источник
Мембранный потенциал

Мембранный потенциал (более точно: трансмембранное напряжение ) представляет собой специальное электрическое напряжение между двумя жидкостными пространствами , в которых заряженных частицы ( ионы ) , присутствуют в различных концентрациях . Мембранный потенциал возникает, когда пространства для текучей среды разделены мембраной, которая позволяет проходить по крайней мере одному из этих типов частиц, но не может проходить одинаково хорошо для всех типов частиц ( полупроницаемость ). Под действием случайного движения частиц большее количество ионов одного типа затем переключается в сторону с более низкой концентрацией, чем в противоположном направлении ( диффузия ), что приводит к разделению зарядов и, таким образом, к трансмембранному напряжению. Разделение зарядов также означает возрастающую силу электрического отталкивания следующих частиц, которая в конечном итоге становится такой же сильной, как и диффузия. В этом состоянии равновесия частицы рассматриваемого типа все еще движутся через мембрану, но в любой момент одинаковое количество в обоих направлениях; чистый ток равен нулю, а мембранный потенциал стабилен. Стабильные мембранные потенциалы также существуют, когда задействованы несколько типов частиц; В этом случае чистый ток для каждого отдельного типа частиц почти всегда не равен нулю, в то время как чистый ток, рассчитанный по всем частицам, также равен нулю.
Мембранные потенциалы имеют первостепенное значение в биологии; все живые клетки создают мембранный потенциал. Чтобы создать и поддерживать разницу в концентрации, они используют молекулярные насосы, такие как натрий-калиевая АТФаза ; избирательная проницаемость создается через определенные ионные каналы . Клетки используют мембранный потенциал для переноса через мембрану ; Мирские вариабельные мембранные потенциалы координировать действия сердечных и направлять и интегрировать информацию в мозге и нервах.
Содержание
Физический состав

Мембранный потенциал представляет собой электрическое напряжение поперек электрохимического двойного слоя , который состоит из мембраны и пространственных зарядов в соседних электролитов . На диаграмме напротив красные и синие области представляют соответственно положительный и отрицательный избыточные заряды . Огибающие кривые представляют плотность заряда, которая круто спадает к центру; это то место, где находится гидрофобная часть мембраны, которой избегают заряды гидратированной оболочки. Плотность заряда экспоненциально падает наружу. Это также относится к напряженности электрического поля (фиолетовая кривая, обычно интеграл по путям плотности заряда), которая полностью исчезает вдали от мембраны, если там не течет ток , так что электрический потенциал (зеленая кривая, интеграл по траекториям напряженности поля) остается там постоянным. является. Разность потенциалов (напряжение) между этими постоянными значениями (зеленая стрелка) называется мембранным потенциалом , а также трансмембранным потенциалом или даже электрическим градиентом , который физики понимают как напряженность поля.
Как и в случае с конденсатором, мембранный потенциал может возникать от внешнего заряда, например, в миелинизированных участках нервных волокон. В биологическом контексте, однако, формирование мембранного потенциала через различие в концентрации по обе стороне мембраны , в связи с ионно-селективной проницаемостью , контролируемой проницаемостью и активным транспортом из ионов через мембрану является более важным.
В случае крупных клеток, таких как нервные или мышечные клетки , мембранный потенциал варьируется в пространстве. Там он используется для передачи и распространения сигналов, а также для обработки информации в сенсорных клетках и центральной нервной системе . В хлоропластах и митохондриях мембранный потенциал служит для энергетической связи процессов энергетического метаболизма : один процесс, см. Цепь переноса электронов , переносит ионы против напряжения и работает , другой, см. АТФ-синтазу , управляется разностью потенциалов.
Измерение

Поскольку потенциал на обеих сторонах мембраны (при условии отсутствия токов) практически не изменяется после небольшого расстояния от мембраны, электрода где-нибудь в двух растворах электролита достаточно для измерения трансмембранного напряжения. Обычно физиологический знак выражается как «внутренний минус внешний потенциал». Для измерения потенциала плазматической мембраны это означает, что один из двух электродов должен быть вставлен в цитозоль ; измеренный мембранный потенциал также может быть интерпретирован как потенциал цитозоля, если внеклеточная жидкость выбрана в качестве точки отсчета.
Измерение мембранных потенциалов на микроскопических структурах, по возможности без электрических, химических и механических помех, затруднено. На фотографии показан вывод внутреннего потенциала ячейки с тонким стеклянным капилляром. При открытии капилляра имеется небольшой диффузионный потенциал, поскольку он заполнен сильным электролитом в высокой концентрации, например B. 3 M KCl, чтобы обеспечить определенный переход к металлическому проводнику, который находится в капилляре и виден на краю изображения. Регистрация внешнего потенциала, который также необходим для измерения мембранного потенциала, на рисунке не изображена.
Физиологические ценности
Фосфолипидный двойной слой с единичной мембраной имеет гидрофобную сердцевину , которая держит заряды пространства друг от друга в хорошем пяти нм . Потенциал покоя клеток животных составляет -70 мВ. Это приводит к напряженности поля более 10 7 В / м, что примерно в четыре раза превышает электрическую прочность воздуха. Электропорация происходит при напряжениях от 0,7 до 1,1 вольт .
Коэффициенты напряженности поля, диэлектрической проницаемости материала мембраны (≥ 2) и постоянной электрического поля приводят к плотности поверхностного заряда почти 3 · 10 −4 Кл / м², что с учетом постоянной Фарадея составляет 3 · 10 −6 мЭкв / м².
Длина Дебая, которая характерна для экспоненциального уменьшения плотности объемного заряда, в физиологических условиях составляет менее одного нанометра. В этом слое есть положительные и отрицательные подвижные носители заряда с поверхностной концентрацией около 2 · 10 -4 мЭкв / м². Таким образом, чистый заряд составляет всего около 1% от плотности заряда.
Изменения потенциала как сигналы
Нервные клетки кодируют информацию в виде кратковременных изменений потенциала. Их можно разделить на две группы, которые имеют разные свойства и функции:
- Градуированные потенциалы проявляются как рецепторный потенциал в сенсорных клетках и как постсинаптический потенциал в химических синапсах и моторной замыкательной пластинке .
- Потенциалы действия генерируются на холме аксона и на аксоне нервной клетки или на подсинаптической мембране мышечных клеток.
Сравнение в обзоре:
| потенциал выпускника | Потенциал действия | |
|---|---|---|
| 1 | амплитудно-модулированный | частотно-модулированный |
| 2 | градуированная амплитуда | постоянная амплитуда |
| 3 | не тугоплавкий | огнеупорный |
| 4-й | Возможно суммирование | суммирование невозможно (принцип «все или ничего») |
| 5 | Пассивное распространение с уменьшением амплитуды | Активное распространение с сохранением амплитуды |
| Шестой | нет порога срабатывания | определенный порог срабатывания |
| 7-е | Деполяризация или гиперполяризация с последующей реполяризацией | только деполяризация с последующей реполяризацией |
| 8-е | неспецифические катионные каналы | быстрые, управляемые по напряжению каналы ионов натрия |
| 9 | Продолжительность от 40 до 4000 мс | Продолжительность 4 мс |
Основы создания
распространение
Растворенные частицы находятся в постоянном движении, что выражается в их тепловой энергии . Это движение называется броуновским молекулярным движением, и оно абсолютно ненаправленное. Если существует связь между двумя местоположениями, которые различаются по концентрации типа частиц, которые могут быть переданы этим типом частиц, чистые частицы перемещаются в направлении более низкой концентрации, пока, наконец, не будут достигнуты одинаковые концентрации. Этот процесс называется диффузией ; он не движется силой, а возникает исключительно в результате случайного движения. Одиночная частица также может двигаться в другом направлении; однако при большом количестве частиц всегда происходит выравнивание концентрации, потому что это вероятно или, другими словами, термодинамически выгодно.
Равновесный потенциал
Когда заряженные частицы ( ионы ) диффундируют, ситуация усложняется, поскольку разделение зарядов связано с диффузией. Это создает возрастающую электростатическую силу, которая ускоряет ионы в направлении более высокой концентрации, так что их движение больше не является полностью ненаправленным и, следовательно, не может быть полной компенсации концентрации. Но и здесь в какой-то момент установится равновесие (состояние без сетевого тока), а именно, когда электрический и стохастический приводы в точности уравновешивают друг друга. Электрическое напряжение , при котором это имеет место, называется равновесным потенциалом и может быть вычислено , используя уравнение Нернста .
Камера , заполненная с солевым раствором будет разделена на две половины клеток с помощью селективной, полупроницаемой мембраны , что только позволяет натрий катиону проходить через него . В каждой полуячейке есть электрод ; два электрода соединены друг с другом с помощью вольтметра, который в этот момент показывает ноль. Если больше поваренной соли растворяется в одной из двух полуячеек (например, правой), сначала наблюдается увеличение напряжения, которое затем остается «равновесным напряжением».
Из-за градиента концентрации ионы натрия диффундируют через мембрану. Из-за разделения зарядов нарастает градиент потенциала: внутренняя часть мембраны (левая камера) становится положительной, внешняя (правая камера) становится отрицательной. Из-за нарастающего потенциального градиента скорость диффузии замедляется. Равновесие диффузии достигается, когда движущая сила градиента концентрации для диффузии влево так же велика, как движущая сила градиента потенциала для диффузии вправо. Чистый заряд, который прошел до этой точки, настолько мал, что концентрации в двух полуячейках существенно не изменились.
Общий мембранный потенциал
Если мембрана проницаема для нескольких ионов, для которых равновесные потенциалы различаются, термодинамическое равновесие невозможно при данных концентрациях . Тем не менее, здесь также устанавливается временно устойчивое состояние: потенциал нулевого тока. При этом напряжении есть токи для отдельных типов ионов, но все токи вместе составляют ноль. Мембранный потенциал ближе к равновесному потенциалу одной из разновидностей ионов, тем выше проницаемость для этой разновидности ионов; точное значение можно рассчитать с помощью уравнения Гольдмана .
Поддержание постоянной концентрации
Ионные токи в случае общего мембранного потенциала уменьшили бы разницу концентраций в долгосрочной перспективе, так что напряжение изменилось бы и в конечном итоге достигло бы нуля. Для постоянного мембранного потенциала необходим механизм, который переносит ионы обратно против направления пассивных токов. Этот транспорт активен , т.е. т.е. ему нужна энергия Гиббса . Натрий-калиевый насос имеет большое значение в биологических системах . Это создает три ионов натрия в обмен на два ионов калия из клетки, энергия Гиббса происходит от гидролиза с АТФ в АДФ и фосфат . Натрий-калиевая АТФаза косвенно управляет дальнейшим обратным транспортом ионов через котранспортеры; это называется вторичным или третичным активным мембранным транспортом .
Получение через свободную энтальпию

По свободной энтальпии Δ G можно определить, движутся ли частицы через мембрану при заданном мембранном потенциале и заданных соотношениях концентраций:
- Если Δ G = 0, существует термодинамическое равновесие : количество частиц, диффундирующих через мембрану за период, одинаково в обоих направлениях.
- Если Δ Gт. Е. Происходит произвольно в рассматриваемом направлении.
- Если Δ G > 0, транспорт является эндергоническим , то есть происходит только в рассматриваемом направлении, если он связан с экзэргоническим процессом (таким как гидролиз АТФ).
Свободную энтальпию также можно рассматривать как меру электрохимического потенциала , возникающего из двух компонентов:
Химический потенциал — без учета разности потенциалов
Формула применяется к транспортировке снаружи внутрь (импорт)
Δ г знак равно Р. ⋅ Т ⋅ пер c ( А. я ) c ( А. а ) <\ displaystyle \ Delta G = R \ cdot T \ cdot \ ln <\ frac  .
.
R : Общая газовая постоянная R = 8,3143 Дж · моль -1 · K -1 T : температура в Кельвинах c (A i ), c (A a ): мольные концентрации вещества A внутри и снаружи ln: натуральный логарифм
Для T = 298 K и с использованием десятичного логарифма уравнение упрощается до
Δ г знак равно 5 , 7-е k J / м О л ⋅ LG c ( А. я ) c ( А. а ) <\ Displaystyle \ Delta G = 5 <,>7 \, \ mathrm <кДж / моль>\ cdot \ lg <\ frac .
- Если концентрация вещества А внутри точно такая же, как снаружи, то Δ G = 0, происходит выравнивание концентрации и перенос вещества не происходит.
- Если концентрация внутри больше, чем снаружи, Δ G > 0, пассивный («добровольный») перенос веществ снаружи внутрь отсутствует.
- Если концентрация снаружи больше, чем внутри, Δ G Электрический потенциал — без учета разницы в концентрации
Свободная энтальпия для переноса чистого заряда равна
Δ г Грузовой транспорт знак равно Z ⋅ Ф. ⋅ Δ Ψ <\ displaystyle \ Delta G _ <\ text <Транспорт заряда>> = Z \ cdot F \ cdot \ Delta \ Psi> .
Z : количество зарядов Z соответствует заряду иона транспортируемой частицы. Он положительный для катионов и отрицательный для анионов. F : постоянная Фарадея F = 96485 Кл · моль -1 ΔΨ: мембранный потенциал
Электрохимический потенциал
Ибо импорт заряженных частиц в результате сложения
Δ г знак равно Р. ⋅ Т ⋅ пер c ( А. я ) c ( А. а ) + Z ⋅ Ф. ⋅ Δ Ψ <\ Displaystyle \ Delta G = R \ cdot T \ cdot \ ln <\ frac .
Уравнение Нернста
В случае равновесия (Δ G = 0) равновесный потенциал ΔΨ 0 для иона может быть определен путем преобразования приведенного выше уравнения:
Δ Ψ 0 знак равно — Р. ⋅ Т Z ⋅ Ф. ⋅ пер c ( А. я ) c ( А. а ) <\ displaystyle \ Delta \ Psi _ <0>= — <\ frac <\ mathrm
для Z = 1 (для Na + , K + ) и T = 298 K использование десятичного логарифма приводит к упрощенному уравнению
Δ Ψ 0 знак равно — 59 , 1 м V ⋅ LG c ( А. я ) c ( А. а ) <\ Displaystyle \ Delta \ Psi _ <0>= — 59 <,>1 \, \ mathrm .
Уравнение Гольдмана-Ходжкина-Каца
Уравнение Гольдмана обеспечивает нулевой потенциал тока при рассмотрении нескольких ионов, оно приведено здесь без вывода:
Δ Ψ знак равно Р. Т Ф. ⋅ пер Σ [ п k а ⋅ c k а — а ] + Σ [ п а п ⋅ c а п — я ] Σ [ п k а ⋅ c k а — я ] + Σ [ п а п ⋅ c а п — а ] <\ Displaystyle \ Delta \ Psi = <\ frac
P : проницаемость каналов для анионов (an) и катионов (ka) c : Концентрация анионов и катионов внутри (-i) или вне (-a) ячейки
Пример мембранного потенциала (смешанный потенциал) ΔΨ, равного -53 мВ при 298 K:
Источник