- Вопрос 71.Структура гена у прокариот. Оперон.
- 1.2. Организация генетического материала
- Читайте также
- Подходы к выявлению чужеродного генетического материала в пищевой продукции
- 1. Организация наследственного материала бактерий
- 8.6. Значение материала патологии для изучения системной организации поведения
- Глава 11. Механика генетического кодирования (XI)
- Глава А. Аналоговые таблицы генетического кода (XIII)
- Глава Б. Барионная оцифровка генетического кода (XIV)
- Глава 11. Механика генетического кодирования (XI)
- Глава A. Аналоговые таблицы генетического кода (XIII)
- Мутационный процесс — первый поставщик эволюционного материала
- Колебания численности — второй поставщик материала для эволюции
- Изучение динамики генетического состава популяции
- Поиски периодической системы. в эволюции
- Немного о Корнях
- Сначала Была Популяция
- Библия Молекулярной Генетики
- Принцип Принципу не помеха
- Букет Эволюционных Тенденций
- Молекулярные Часы Эволюции
- Конструктор для Создателя
Вопрос 71.Структура гена у прокариот. Оперон.
Ген определяют как участок молекулы ДНК (у некоторых вирусов РНК), кодирующий первичную структуру полипептида, молекулы транспортной или рибосомной РНК либо взаимодействующий с регуляторным белком.
Ген — это последовательность нуклеотидов, которая выполняет определенную функцию в организме, например последовательность нуклеотидов , кодирующая полипептид тРНК или обеспечивающая транскрипцию другого гена.
Прокариоты – это организмы, в клетках которых отсутствует оформленное ядро. Его функции выполняет нуклеоид (то есть «подобный ядру»); в отличие от ядра, нуклеоид не имеет собственной оболочки.
Тело прокариот, как правило, состоит из одной клетки. Однако при неполном расхождении делящихся клеток возникают нитчатые, колониальные и полинуклеоидные формы (бактероиды). В прокариотических клетках отсутствуют постоянные двумембранные и одномембранные органоиды: пластиды и митохондрии, эндоплазматическая сеть, аппарат Гольджи и их производные. Их функции выполняют мезосомы – складки плазматической мембраны. В цитоплазме фотоавтотрофных прокариот имеются разнообразные мембранные структуры, на которых протекают реакции фотосинтеза.
Размеры прокариотических клеток изменяются от 0,1-0,15 мкм (микоплазмы) до 30 мкм и более. Большинство бактерий имеет размеры 0,2-10 мкм. У подвижных бактерий имеются жгутики, основой которых служит белки флагеллины.
Структура гена прокариот проста. Участок , кодирующий определенный белок, представляет ряд нуклеотидов (триплетных кодонов), которые транскрибируются на мРНК и затем транслируются на рибосоме в данный белок. Более сложной является система регуляции синтеза белка у бактерий. Как показали исследования, проведенные на E.coli, структурные гены, детерминирующие утилизацию этой бактерии лактозы, довольно тесно сцеплены и образуют оперон.
Оперон – участок бактериальной хромосомы, включающий следующие участки ДНК: Р – промотор, О – оператор, Z, Y, А – структурные гены, Т – терминатор. (В состав других оперонов может входить до 10 структурных генов.)
Промотор служит для присоединения РНК-полимеразы к молекуле ДНК с помощью комплекса CAP-цАМФ (CAP – специфический белок; в свободной форме является неактивным активатором; цАМФ – циклоаденозинмонофосфат – циклическая форма аденозинмонофосфорной кислоты).
Оператор способен присоединять белок–репрессор (который кодируется соответствующим геном). Если репрессор присоединен к оператору, то РНК-полимераза не может двигаться вдоль молекулы ДНК и синтезировать иРНК.
Структурные гены кодируют три фермента, необходимые для расщепления лактозы (молочного сахара) на глюкозу и галактозу. Молочный сахар лактоза – менее ценный продукт питания, чем глюкоза, поэтому в присутствии глюкозы сбраживание лактозы является невыгодным для бактерии процессом. Однако при отсутствии глюкозы бактерия вынуждена переходить на питание лактозой, для чего синтезирует соответствующие ферменты Z, Y, А.
Терминатор служит для отсоединения РНК-полимеразы после окончания синтеза иРНК, соответствующей ферментам Z, Y, А, необходимым для усвоения лактозы.
Для регуляции работы оперона необходимы еще два гена: ген, кодирующий белок–репрессор, и ген, кодирующий белок СYА. Белок СYА катализирует образование цАМФ из АТФ. Если в клетке имеется глюкоза, то белок СYА вступает с ней в реакцию и переходит в неактивную форму. Таким образом, глюкоза блокирует синтез цАМФ и делает невозможным присоединение РНК-полимеразы к промотору. Итак, глюкоза является репрессором.
Если же в клетке имеется лактоза, то она взаимодействует с белком–репрессором и превращает его в неактивную форму. Белок–репрессор, связанный с лактозой, не может присоединиться к оператору и не преграждает путь РНК-полимеразе. Итак, лактоза является индуктором.
Предположим, что первоначально в клетке имеется только глюкоза. Тогда белок–репрессор присоединен к оператору, а РНК-полимераза не может присоединиться к промотору. Оперон не работает, структурные гены выключены.
При появлении в клетке лактозы и при наличии глюкозы белок–репрессор отщепляется от оператора и открывает путь РНК-полимеразе. Однако РНК-полимераза не может присоединиться к промотору, поскольку глюкоза блокирует синтез цАМФ. Оперон по-прежнему не работает, структурные гены выключены.
Если же в клетке имеется только лактоза, то белок–репрессор связывается с лактозой, отщепляется и открывает путь РНК-полимеразе. В отсутствии глюкозы белок СYА катализирует синтез цАМФ, и РНК-полимераза присоединяется к промотору. Структурные гены включаются, РНК-полимераза синтезирует иРНК, с которой транслируются ферменты, обеспечивающие сбраживание лактозы.
Таким образом, лактозный оперон находится под двойным контролем индуктора (лактозы) и репрессора (глюкозы).
Белок laс –репрессор –аллостерический белок, который имеет два центра связывания и поэтому может присоединяться к двум разным молекулам. В одном случае с оператором, а в другом- с лактозой.Это кластерная организация генов.
Источник
1.2. Организация генетического материала
1.2. Организация генетического материала
Структурно-функциональная организация генетического аппарата определяет деление всех живых организмов на прокариот и эукариот. У прокариот (к которым относятся бактерии и археи) ДНК представлена кольцевой молекулой и находится в цитоплазме клетки. У эукариот (к которым относятся все остальные организмы) структурными носителями генетической информации ДНК являются хромосомы, находящиеся в ядре.
Хромосомы представляют собой сложную многоуровневую структуру, в которой ДНК взаимодействует с различными белками. Базовым уровнем этой структуры являются нуклеосомы, представляющие собой глобулы из восьми молекул белков гистонов, обвитых ДНК. Нуклеогистоновая нить в дальнейшем многократно укладывается, формируя компактные хромосомы. Такая структура открывает широкие возможности для регуляции.
Поскольку число генов в организме несоизмеримо больше числа хромосом, то понятно, что каждая хромосома несет много генов. Каждый ген занимает в хромосоме определенное место – локус. Гены, расположенные на одной хромосоме, называются сцепленными.
Помимо ядра, небольшая доля генетической информации эукариотической клетки находится в таких органоидах, как митохондрии и хлоропласты, имеющих собственные генетические системы: свои ДНК, разнообразные РНК (и-РНК, т-РНК, р-РНК) и рибосомы, что позволяет осуществлять независимый синтез белка. Кольцевые ДНК этих органоидов явились важным доводом в пользу их бактериального симбиотического происхождения на заре формирования жизни.
Клеточное ядро эукариот разделяет процессы транскрипции и трансляции, что предоставляет широкие возможности для регуляции. Регуляция происходит на всех этапах экспрессии генов эукариот. Дополнительным этапом у них является процессинг – процесс сложных преобразований синтезированной в ходе транскрипции РНК. Важнейшая составляющая процессинга и-РНК – сплайсинг, при котором происходит вырезание интронов (некодирующих областей гена) и сшивание экзонов (кодирующих областей). Экзоны и интроны обусловливают «мозаичную» структуру эукариотических генов. Именно в результате процессинга синтезированная в ядре РНК становится функционально активной.
Понимание многообразных механизмов регуляции послужило причиной радикальных изменений наших представлений о структурно-функциональной организации генетического аппарата в настоящее время.
Одним из основателей современной генетики, выдающимся датским ученым В. Иоганнсеном (1857–1927) были предложены базовые генетические термины – ген, аллель, генотип, фенотип, определяющие генетические характеристики особи.
Гены, расположенные в своих локусах, могут иметь варианты – аллели. Локус, имеющий в популяции более одного аллеля, называется полиморфным. Обычно аллели обозначаются буквами латинского или греческого алфавита, а если их много, то с верхним индексом. Количество аллелей разных генов в популяциях организмов может быть различным. Одни гены имеют много аллелей, другие – мало. В любом случае число аллелей ограничено эволюционными факторами: аллели, ухудшающие адаптивные свойства вида либо несовместимые с жизнью, элиминируются естественным отбором.
Конкретный организм эукариот имеет только два аллеля одного гена: по числу гомологичных локусов гомологичных хромосом (отцовской и материнской). Организм, у которого оба аллеля одинаковые, называется гомозиготой (по данному гену). Организм, у которого аллели разные, называется гетерозиготой (рис. 1.4). Аллели, локализованные на половых хромосомах гетерогаметного пола, могут присутствовать в единственном числе.
Генотип можно представить как совокупность аллелей организма, а фенотип – как совокупность его внешних признаков.
Введенный в 1920 г. немецким ботаником Г. Винклером (1877–1945) термин геном стал характеристикой целого вида организмов, а не конкретной особи. Это понятие в дальнейшем стало одним из важнейших. К 1980-м гг. XX в. формируется новое направление генетики – геномика. Первоначально геном характеризовали как совокупность генных локусов гаплоидного набора. Однако оказалось, что сами гены занимают относительно небольшую часть генома, хотя и составляют его основу. Большую часть занимают межгенные участки, где есть области с регуляторной функцией, а также районы невыясненного пока назначения. Регуляторные участки неразрывно связаны с генами, являются своего рода «инструкциями», определяющими работу генов на разных этапах развития организма. Поэтому геномом в настоящее время называют всю совокупность ДНК клетки, характерную для ДНК вида.
На современном этапе развития генетики геномика становится одним из ее ключевых разделов. Успехи геномики наглядно продемонстрировало успешное завершение программы «Геном человека».
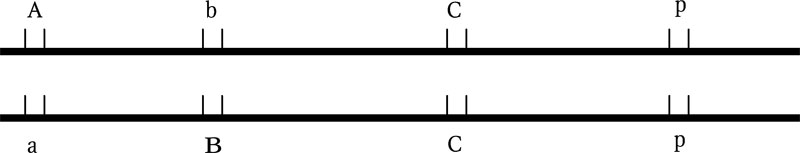
Рис. 1.4. Аллели сцепленных генов двух гомологичных хромосом
Данный текст является ознакомительным фрагментом.
Продолжение на ЛитРес
Читайте также
Подходы к выявлению чужеродного генетического материала в пищевой продукции
Подходы к выявлению чужеродного генетического материала в пищевой продукции Требование по маркировке продуктов, содержащих ГМ сою или кукурузу, «кроме случаев, когда отсутствуют генетически модифицированные белки или ДНК. » (Положение ЕС 1139/98) привело к развитию
1. Организация наследственного материала бактерий
1. Организация наследственного материала бактерий Наследственный аппарат бактерий представлен одной хромосомой, которая представляет собой молекулу ДНК, она спирализована и свернута в кольцо. Это кольцо в одной точке прикреплено к цитоплазматической мембране. На
8.6. Значение материала патологии для изучения системной организации поведения
8.6. Значение материала патологии для изучения системной организации поведения Таким образом, проекция индивидуального опыта на структуры мозга изменяется в филогенезе, определяется историей обучения в процессе индивидуального развития и модифицируется при
Глава 11. Механика генетического кодирования (XI)
Глава 11. Механика генетического кодирования (XI) Об этом можно прочитать в любом учебнике. И все же — чтобы облегчить понимание последующих рассуждений — позволим себе очень коротко остановиться на работе машины кодирования. Барбьери связывает формирование таких машин с
Глава А. Аналоговые таблицы генетического кода (XIII)
Глава А. Аналоговые таблицы генетического кода (XIII) Первым, кто попытался упорядочить таблицу генетического кода и построить ее на рациональной основе, был наш выдающийся ученый Юрий Борисович Румер. Он был физиком, учеником Макса Борна, хорошо знал Альберта Эйнштейна,
Глава Б. Барионная оцифровка генетического кода (XIV)
Глава Б. Барионная оцифровка генетического кода (XIV) ФОРМАТЫ 1D и 2D Строго говоря, барионным числом называется сохраняемое квантовое число системы. Нам нет необходимости углубляться в эту тему. Может быть, стоит помнить лишь то, что барион — это элементарная частица,
Глава 11. Механика генетического кодирования (XI)
Глава 11. Механика генетического кодирования (XI) Об этом можно прочитать в любом учебнике. И все же – чтобы облегчить понимание последующих рассуждений – позволим себе очень коротко остановиться на работе машины кодирования. Барбьери связывает формирование таких
Глава A. Аналоговые таблицы генетического кода (XIII)
Глава A. Аналоговые таблицы генетического кода (XIII) Первым, кто попытался упорядочить таблицу генетического кода и построить ее на рациональной основе, был наш выдающийся ученый Юрий Борисович Румер. Он был физиком, учеником Макса Борна, хорошо знал Альберта Эйнштейна,
Мутационный процесс — первый поставщик эволюционного материала
Мутационный процесс — первый поставщик эволюционного материала Элементарные эволюционные факторы выделяют на основе характера и природы их воздействия на популяции, а также по результатам оказываемого ими давления на популяции. При этом необходимым и достаточным
Колебания численности — второй поставщик материала для эволюции
Колебания численности — второй поставщик материала для эволюции Один из важнейших эволюционных факторов — периодические изменения численности особей, популяционные волны. В данном случае речь идет о колебаниях в положительную и отрицательную сторону, сменяющих друг
Изучение динамики генетического состава популяции
Изучение динамики генетического состава популяции В начале этой книги подчеркивалось, что одна из важнейших задач современного популяционного исследования — получение материалов по самым разнообразным эволюционным ситуациям в природных популяциях, в частности,
Источник
Поиски периодической системы. в эволюции
За последние сто лет были выявлены универсальные генетические конструкции, лежащие в основе жизнедеятельности самых разных живых существ. Но как с этим принципом уживается бесконечное многообразие и специфичность существующих видов?
Биологии, бесспорно, не суждено в обозримом будущем приобрести гордый статус «точной». В этом смысле из всех биологических дисциплин наибольшие шансы имеют генетика и молекулярная биология. Именно родоначальник генетики Грегор Мендель впервые в биологии использовал методы математического анализа. А его последователи открыли основные пути и принципы передачи наследственной информации в череде поколений, переведя расплывчатые биологические формулировки на строгий язык химических понятий.
Стремительное накопление знаний о строении геномов и структуры генов у разных организмов позволило современным «Менделеевым», точно классифицировав элементы генетического «конструктора», делать выводы о закономерностях в организации наследственного материала живого.
Немного о Корнях
Кстати сказать, со времен Чарльза Дарвина никто так и не придумал хорошей альтернативной теории эволюции, несмотря на частые заявления об обратном. Фундаментальный принцип естественного отбора до сих пор никем не опровергнут. А вот что касается более тонких механизмов. По большому счету, наши представления об эволюции являются всего лишь зеркалом, отражающим полноту наших знаний о структуре и функционировании биологических систем. И не вина зеркала, если лицо, скажем мягко, несколько «подкачало». Это относится и к удельному весу различных механизмов, принимающих участие в эволюционном процессе, начиная от горизонтального переноса генов и т. д.
Нужно заметить, что поиски основополагающих эволюционных принципов ранее проводились исключительно на морфологическом уровне, сейчас же акцент все более смещается на уровень молекулярный. Давайте заглянем ненадолго в прошлое, чтобы лучше представить себе «эволюцию» наших представлений об эволюции.
Сначала Была Популяция
В основе современной синтетической теории эволюции лежат достижения генетики. Однако в начале XX столетия первые адепты новой науки сами были «махровыми» антидарвинистами, утверждавшими, что новые виды рождаются исключительно благодаря мутационному процессу без всякого участия естественного отбора. Так продолжалось до 1908 года, когда английский математик Г. Харди и немецкий врач В. Вайнберг независимо сформулировали знаменитый закон, объяснивший наблюдаемое в природе сохранение изменчивости при передаче признаков от родителей к потомкам. Это событие ознаменовало рождение наиболее формализованного направления в биологии — популяционной генетики.
Доминирующей в настоящее время является синтетическая теория эволюции или ее более модернизированные производные. И хотя основой теории считаются достижения генетики, «любовь», тем не менее, здесь возникла не с «первого взгляда»
Закон Харди–Вайнберга представляет собой довольно простую математическую формулу, согласно которой при постоянных условиях имеющаяся в популяции изменчивость будет оставаться на одном и том же уровне. Но в природе редко встречаются равновесные популяции: под действием различных внешних факторов в популяции происходит изменение первоначальных частот
(концентрации) генов. И вот это уже — элементарное микроэволюционное событие. К слову, положение о том, что единицей эволюции является не особь, а популяция, сформулировал «зубр» русской генетики Н. Тимофеев-Ресовский, а сами термины макроэволюция и микроэволюция ввел Ю. Филипченко, читавший в 1913 году первый в России университетский курс «Учение о наследственности и эволюции».
Наши соотечественники Н. Дубинин и Д. Ромашов впервые заговорили о генетико-автоматических процессах в популяциях, т. е. о микроэволюционных событиях, не связанных напрямую с естественным отбором. Это значит, что, кроме направленного изменения популяции под действием отбора, частоты генов могут меняться достаточно случайным образом под действием таких факторов, как резкие изменения численности («волны жизни»), соотношения полов и т. п. Интересно, что процесс случайного генетического дрейфа ученые смоделировали с помощью «тиражей» с разноцветными шариками, изображавшими различные генные вариации. Эти представления были предтечей будущей теории нейтрализма, которая, как казалось, потрясла основы теории эволюции. Но об этом — позже.
Библия Молекулярной Генетики
А теперь пройдем снова тот же исторический путь, но уже по следу формирования представлений о гене как элементарной единице генетической информации.
Все началось с менделевских наследственных факторов, названных в 1909 году датчанином В. Иогансеном генами. В работах школы Т. Моргана было установлено, что ген, являющийся частью хромосомы, мутирует как целостная структура и контролирует определенный признак организма. Благодаря открытию влияния ионизирующего излучения на мутационный процесс ученые получили инструмент для изучения протяженности, структуры и делимости самого гена, и первенство здесь принадлежит А. Серебровскому, основателю кафедры генетики Московского университета.
Далее Дж. Бидл и Э. Тейтем выдвинули предположение, что один ген кодирует синтез одного фермента. А затем, в исследованиях Дж. Уотсона и Ф. Крика, произошла идентификация гена как участка молекулы ДНК, завершившаяся расшифровкой свойств самого генетического кода. Французские ученые Ф. Жакоб и Ж. Моно открыли явление генетического контроля синтеза ферментов у бактерий, позволившее увидеть реальный механизм регуляции активности генов. С. Бензер и Ч. Яновский установили, что наименьшей изменяемой единицей в пределах гена является пара нуклеотидов. И в этой точке мы от биологических переходим уже к чисто химическим понятиям.
Я готов подвергнуться некоторым нападкам, тем не менее утверждаю, что не существует «специальных» механизмов эволюции
В итоге сформировалась центральная догма молекулярной биологии, наиболее полно отражающая представления о структуре и функционировании гена. Вот она: поток информации, записанной в ДНК, идет от ДНК к информационной РНК, на основе которой затем синтезируется белковая молекула. И обратите внимание — этот поток является дорогой с односторонним движением: информация, попавшая в белок, оттуда не возвращается! Не может быть синтезирована кодирующая последовательность ДНК на основе белковой последовательности аминокислот, что является подтверждением закона о невозможности наследования благоприобретенных признаков.
Принцип Принципу не помеха
Перечисленные выдающиеся результаты породили некую иллюзию, что все и у всех построено одинаково. И это казалось довольно справедливым, поскольку на самом деле пути метаболизма у всех живых существ в принципе очень схожи. Например, синтез какого-нибудь нуклеотида одинаков — что у дрожжей, что у человека. Рибосомы — эти цеха по производству белка — устроены практически одинаково у разных организмов. Синтез информационной РНК — тоже универсальный механизм.
Внимательно приглядевшись, мы обнаруживаем скорее талантливые вариации на заданную тему, чем принципиально новые эволюционные приобретения. Возникает закономерное подозрение, что существуют некоторые универсальные конструкции, которые «подтверждаются» на ранних стадиях эволюции, а затем тиражируются и используются самыми разными организмами. Прекрасная иллюстрация одного из основополагающих принципов биологии — принципа биологической универсальности!
Но, как и положено по законам философии, этот принцип уравновешивается и дополняется принципом биологической специфичности. Например, было установлено, что сама организация наследственного материала и структура генов значительно различаются у разных организмов. Естественным образом рождается вопрос: можно ли выделить какие-то универсальные блоки, найти закономерности в этом генетическом разнообразии живых существ?
Букет Эволюционных Тенденций
Попробуем выявить эти принципиальные отличия между организмами, сравнивая геномы прокариот (одноклеточных организмов, не имеющих клеточного ядра) и эукариот, у которых наследственный материал содержится в полноценном клеточном ядре, покрытом оболочкой. Считается, что более «продвинутые» эукариоты произошли от прокариот и «моложе» их примерно на 500 млн лет.
Что же мы видим? Для бактерий, принадлежащих к прокариотам, характерна оперонная организация генома. Благодаря этому качеству бактерии способны быстро переключать метаболизм с одного питательного вещества на другой, что помогает им оперативно реагировать на изменения в окружающей среде. При этом структурные гены, кодирующие ферменты определенного пути биосинтеза, объединены у бактерий в оперон — группу, имеющую общий регуляторный элемент. На такой матрице синтезируется единая молекула информационной РНК, на которой закодированы ферменты всех этапов биосинтеза. А вот у эукариот, включая сюда и грибы, опероны не обнаружены, и те же самые структурные гены у них могут быть разбросаны по разным хромосомам. Таким образом, можно выделить первую тенденцию в эволюции ге-нома — тенденцию к автономизации генов.
Формирование молекулярной парадигмы в 60-е годы прошлого века породило иллюзию, наиболее ярко отразившуюся во
фразе-каламбуре Жака Моно: “What is true for E. coli, is true for E. Lephant” («Что справедливо для кишечной палочки, справедливо и для слона») В кажущееся противоречие с этой тенденцией вступает другая — стремление к олигомеризации генома. Что это значит? Только то, что некоторые гены, ранее существовавшие по отдельности, сливаются в единый ген. А закодированная на нем белковая молекула служит ферментом сразу в нескольких метаболических реакциях. И по мере усложнения биологической организации, от грибов к дрозофиле и человеку, число таких метаболических реакций, осуществляемых одним белком-ферментом, возрастает.
Следующая интересная тенденция — возникновение мозаичной структуры гена. Так, у эукариот сам структурный ген состоит из разных участков — интронов и экзонов. Интроны не являются кодирующими последовательностями и не несут генетической информации. На таком расщепленном гене образуется соответствующая молекула РНК, затем участки, соответствующие интронам, вырезаются, а оставшиеся кусочки — экзоны — сшиваются вместе. И только эта РНК становится матрицей для синтеза белковой молекулы.
У паразитических созданий появляется еще одно удивительное свойство — перекрывание генов. Вирусы, например, имеют очень маленькие геномы, но большие информационные запросы. И они нашли выход из трудного положения: гены у них расположены на манер русской матрешки — один в другом.
Теперь, разобравшись с основными тенденциями в организации генетического материала, переходим к более глобальным обобщениям. Вспомним, что мы живем в эпоху геномики и в настоящее время уже расшифрованы геномы многих организмов. Сегодня мы можем заметить то, о чем лишь интуитивно догадывались: в ходе эволюции число генов увеличивается! Каким же образом возникают новые гены и, соответственно, новые белки и функции?
Молекулярные Часы Эволюции
Проблема эта довольно сложна, и ее не разрешить одними только горизонтальными генетическими переносами, хотя они, несомненно, тоже «имеют место быть». Популяционная генетика доказала, что возникновение крупных новшеств невозможно объяснить простыми мутациями, как нельзя одними точечными мутациями объяснить происхождение человека от обезьяны. На этот процесс просто не хватит времени — к этому выводу давно пришли умные священники.
Из-за простых точечных мутаций в популяции возникает полиморфизм, т. е. разнообразие вариантов одного гена. Согласно теории нейтрализма японца М. Кимуры, в популяции могут сохраняться только те мутации, которые функционально не портят белки (нейтральные мутации). И действительно, до 10 % нуклеотидных по-следовательностей гена и соответствующих аминокислотных последовательностей белка испытывают такие нейтральные замены без видимого ущерба для работы активных центров белковой молекулы.
На основе этой идеи Э. Цукеркандл и Л. Полинг придумали остроумную и практичную вещь: молекулярные часы эволюции. Если на самом деле происходит закрепление нейтральных мутаций, а сам мутационный процесс идет с постоянной скоростью, то по количеству нейтральных замен можно посчитать эволюционное время. К сожалению, в бочке меда, как водится, не обошлось без ложки дегтя: выяснилось, что разные гены накапливают нейтральные мутации с неодинаковой скоростью, а эволюционные деревья, построенные на их основе, могут представлять собой довольно разнокалиберную рощу. Так что без традиционного изучения геологических слоев нам все равно не обойтись.
Конструктор для Создателя
Но мы немного отвлеклись. Понятно, что за счет точечных мутаций эволюция не происходит. Какова же тогда движущая сила эволюционного процесса, а более конкретно — как в природе появляется нечто принципиально новое?
Блочный или блочно-модульный принцип в теории эволюции подразумевает, что все ныне сущее возникло за счет исходно небольшого числа неких инвариантных доменов, которые далее комбинировались в разных сочетаниях
Японец С. Оно предложил простой и понятный механизм: эволюция происходит за счет тиражирования существующих генетических структур и их дальнейшей дивергенции (расхождения). При этом даже неважно, откуда в геноме появляются новые копии генов: с помощью мигрирующих генетических элементов или специальных ферментов, дуплицирующих (удваивающих) участки ДНК. Главное, что существует реальная физическая возможность для умножения как отдельных участков и целых хромосом, так и всего генома.
Вообще биология — это наука, где полно исключений. Там есть всё, вопрос только в том, какой удельный вес имеют те или иные феномены А что потом? Да что угодно! Часть копий может быть утеряна, а часть — сохранена; из разных копий могут потеряться разные гены. Между прочим, если у вас есть две копии одного гена, то одна из них может работать, а другая при этом — беспрепятственно мутировать, как ей вздумается. Эта последняя благополучно ускользает из-под давления естественного отбора и начинает с большой скоростью и без помех накапливать мутации. Так появляются функционально неактивные псевдогены. Но не думайте, что такие псевдогены становятся просто мусорной корзиной: их генетический материал может очень пригодиться для создания новых генов. В принципе, время от времени и сами псевдогены способны активироваться, чтобы предстать перед судом естественного отбора.
Все высказанные соображения приводят нас к довольно простому и ясному выводу, назовем его блочный принцип теории эволюции: все существующее генетическое разнообразие произошло за счет тиражирования и комбинаторики относительно небольшого числа исходных генетических структур. Такой, знаете ли, генетический конструктор, в который наш создатель (не написать ли это слово с прописной буквы?) с успехом играет вот уже многие миллионы лет…
Источник